Grundlagen: Über Zufall, Mutation und die Grenzen der Vererbungslehre
Wie wahrscheinlich ist es, dass sich zehntausende Buchstaben in genau der richtigen Reihenfolge "durch Zufall" aufreihen? Im Grunde ist diese Frage irrelevant, sie lässt sich aber als einfaches mathematisches Problem lösen.
Im Sinne der modernen Evolutionstheorie ist sie freilich sogar dumm gestellt. Denn sie geht davon aus, dass alle Buchstaben zum gleichen Zeitpunkt "auftauchten", nur in einer Anordnung "funktionieren" und nur in einem kulmulierten Moment, nicht nach einander oder in bestimmten vordefinierten Gesetzmäßigkeiten, aufgereiht werden.
Die viel wichtigere Frage ist, was notwendig ist, um eine Veränderung der Buchstabenkombinationen zu erreichen und wie lange es durchschnittlich dauert, bis es eine neue Buchstabenkombination gibt. Und erst dann ist zu überlegen, wie wahrscheinlich es ist, dass in einer bestimmten Zeit, bei einem rein zufälligen Verändern der Kombination, eine ganz bestimmte Buchstabenkombination entsteht.
Genau das, was die Evolutionstheorie beschreibt. Man muss dafür natürlich berücksichtigen, das es kein vordefiniertes Ziel gibt, der aktuelle Ist-Stand also auch hätte völlig anders sein können. Auch ist zu bemerken, dass der Evolution mit ungefähr 3 1/2 - 4 Milliarden Jahren ein unvorstellbar langer Zeitraum zur Verfügung stand und, dass es nicht lediglich ein Individuum gab, das hätte mutieren können.
Aus den Beobachtungen erschließt sich, dass die absolut überwiegende Anzahl an Mutationen entweder überhaupt keinen Effekt hat oder - und das ist schon wesentlich seltener - das Individuum nicht lebensfähig oder zumindest nicht lebensfähiger macht, als andere Individuen. Solche Mutationen werden durch eine gleichbleibende oder schlechtere Populationsrate ausgesiebt, also selektiert.
Bei der riesigen Anzahl an möglichen Mutationen und Individuen, die mutieren können, muss es auch Mutationen geben, die einem Individuum nützen. Das ist ebenfalls wieder nur eine Frage der Wahrscheinlichkeit. Diese Individuen haben durch eine höhere "Fitness" eine höhere Chance, ihr Erbgut in die nächste Generation weiterzugeben. Nützliche Mutationen werden also mit einer höheren Wahrscheinlichkeit weitergegeben. So gibt es in "kurzer" Zeit mehr Individuen, die bereits über diese Mutation verfügen.
Was allerdings nützlich ist, also die "Fitness" erhöht, bestimmt sich nicht objektiv, sondern allein danach, ob es einem Individuum in einer bestimmten Umwelt hilft oder nicht. In einer anderen Umwelt kann die gleiche Mutation fatal sein, was dazu führt, dass sie sich dort nicht durchsetzen kann. Und weil nützliche Mutationen relativ selten auftreten, ist Evolution ein eher langsamer Prozess, der nicht in den allerseltesten Fällen live beobachtet werden kann. Deshalb sehen Menschen, zum Beispiel, im Großen und Ganzen heute noch so aus, wie vor zehntausend Jahren. Aber ein paar tausend Jahre sind auch nichts im Vergleich zu 3 - 4 Milliarden Jahren.
Immer wieder liest man darüber hinaus ja auch - und im von mir rezensierten Video "Hat die Bibel doch Recht? - Der Evolutionstheorie fehlen die Beweise" wird es ebenfalls behauptet - dass es einen Widerspruch zwischen Darwin und Mendel gäbe.
Mendel untersuchte die reine Vererbung - ohne Mutation - von einer Generation auf die andere. Ausschlaggebend hierfür ist die Meiose.
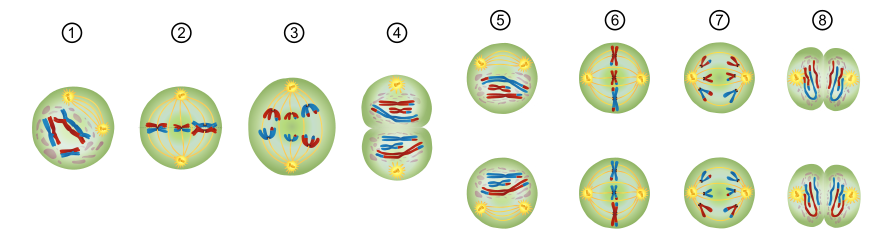
Schema der Meiose. In diesem Beispiel sind drei Chromosomenpaare dargestellt und die Chromosomen je blau bzw. rot gekennzeichnet nach dem Elternteil, von dem sie geerbt wurden. Außerdem sind Mikrotubuli und Centrosomen (beide gelb-orange) dargestellt, um die Phasen der Teilungen besser unterscheiden zu können. Auf (1) Prophase I (hier dargestellt in der Unterphase der Diakinese), (2) Metaphase I, (3) Anaphase I, (4) Telophase I der ersten meiotischen Teilung folgt – nach einer hier nicht dargestellten Zwischenphase der Interkinese – die zweite meiotische Teilung mit (5) Prophase II, (6) Metaphase II, (7) Anaphase II, (8) Telophase II. Vier gleich große Zellen wie hier angedeutet entstehen beispielsweise bei der Spermatogenese, nicht aber bei der Oogenese.
Meiose (griech. "Verminderung", "Verkleinerung"), auch Reifeteilung genannt, ist eine besondere Art Kernteilung eukaryotischer Zellen (#1), die in zwei Schritten abläuft: Meiose I (#2) und Meiose II (#3). Dabei wird die Anzahl der Chromosomen halbiert und es entstehen genetisch voneinander verschiedene Zellkerne. Damit unterscheidet sich die Meiose grundlegend von der sonst gewöhnlichen Kernteilung, der Mitose. Diese lässt den Chromosomenbestand unverändert und bringt damit genetisch identische Zellkerne hervor.
Bei der Meiose wird aus einem doppelten di-poliden Chromosomensatz einen einfachen ha-poliden Chromosomensatz gemacht. Dieser findet sich in den Keimzellen der Eltern wieder, welche bei der Befruchtung miteinander verschmelzen. Erst dadurch wird ein neuer doppelter ha-polider Chromosomensatz geschmiedet, der dann zu einem doppelten di-poliden Chromomensatz ergänzt wird. Ausführlicher ist das alles in den Fußnoten 2 und 3 lesbar. Wenn das alles funktioniert, können dabei natürlich nur die Erbinformationen weitergegeben werden, die bisher in der Elterngeneration auch schon vorhanden waren. Das entspricht Mendels Lehre.
Und das sind ihre Grenzen. Sie stellt den Normalfall dar und gilt da nicht mehr, wo die Erbinformation verändert wird. Dies kann beispielsweise durch einen Fehler bei der Meiose oder aber auch durch Mutation geschehen. Es handelt sich hierbei also nicht um eine nach den mendelschen Regeln vererbte Eigenschaft und nur die Evolutionstheorie gibt weiteren Aufschluss.
Mendel bietet also lediglich die Möglichkeit der Erklärung, wie Merkmale von einer Generation in die nächste getragen werden. Die Vererbungslehre ist also sehr wohl Teil der synthetischen Evolutionstheorie und widerspricht ihr nicht.
#1 - Eukaryotischer Zellen
Eukaryoten oder Eukaryonten (Eukaryota) sind eine Domäne der Lebewesen, deren Zellen (Eucyten) einen echten Kern und eine reiche Kompartimentierung haben. Hierin unterscheiden sie sich von den beiden übrigen Domänen im System der Lebewesen, den prokaryotischen Bakterien und Archaeen mit procytischen Zellen.
Demnach sind Tiere, Pflanzen, Pilze, Algen, einige Bakterien und auch der Mensch Eukaryoten oder Eukaryonten.
#2 - Meiose I (Reduktionsteilung)
Prophase I
Die erste meiotische Teilung beginnt mit der Prophase I. Diese wird in fünf Stadien unterteilt:
Metaphase I und Anaphase I
In der Metaphase I versammeln sich die gepaarten Chromosomen in der Äquatorialebene des Spindelapparats. Auch in dieser Phase können im Lichtmikroskop die Chiasmata sichtbar werden. In der anschließenden Anaphase I werden im Gegensatz zur mitotischen Anaphase nicht einzelne Chromatiden, sondern Chromatidenpaare zu den beiden Spindelpolen bewegt. Auf Grund des vorangegangenen Cross-overs sind die beiden zusammenhängenden Chromatiden jedoch nicht mehr identisch.
Telophase I
In der Telophase I liegt an jedem Pol dann jeweils nur noch ein Chromosom (mit zwei Chromatiden) jedes Typs vor. Es ist also zu einer Reduktion der Chromosomenzahl gekommen. Wie bei der mitotischen Telophase dekondensieren die Chromosomen nun und die Kernhülle bildet sich wieder. An die abgeschlossene Kernteilung schließt sich eine Zellteilung an. Der anschließende Zeitraum bis zum Beginn der zweiten mitotischen Teilung wird als Interkinese bezeichnet.
Bei einem menschlichen euploiden Chromosomensatz enthält der Zellkern der diploiden Zelle vor der meiotischen Teilung 23 Paare duplizierter Chromosomen, also 46 Chromosomen bzw. 92 Chromatiden. Nach der Meiose I hat jeder der beiden Tochterkerne 23 Chromosomen erhalten, die je aus einem Chromatidenpaar bestehen, also 46 Chromatiden.
#3 Meiose II (Äquationsteilung)
Im Anschluss an die Interkinese folgt die Meiose II. Sie ähnelt vom Ablauf her einer mitotischen Teilung, mit dem Unterschied, dass nur noch halb so viele Chromosomen vorliegen.
Prophase II
Die Chromosomen werden erneut kondensiert.
Metaphase II
Die noch aus zwei Chromatiden bestehenden Chromosomen werden in der Äquatorialebene angeordnet und am Centromer getrennt.
Anaphase II
Die getrennten Chromosomen werden einzeln den Tochterkernen zugeteilt.
Telophase II
Die Aufteilung ist abgeschlossen und die Kernhüllen bilden sich wieder aus.
Die aus der Telophase II hervorgehenden Zellkerne enthalten damit jeweils einen haploiden, unverdoppelten Chromosomensatz. Somit sind die vier Chromatiden einer jeden Tetrade der Prophase I jeweils zufällig auf vier verschiedene haploide Zellkerne verteilt worden.
Beide Vorgänge, Meiose I und Meiose II, zusammen bewirken eine effektive Durchmischung des Erbguts der beiden Eltern. Diese Rekombination erhöht die Zahl der genetischen Kombinationen innerhalb der Population und damit die Chance, dass in einigen Individuen für das Überleben günstige Merkmale zusammenkommen und so zu einem Selektionsvorteil führen. Im Vergleich zu asexuellen Organismen ermöglicht dies eine schnellere Evolution.
Wie wahrscheinlich ist es, dass sich zehntausende Buchstaben in genau der richtigen Reihenfolge "durch Zufall" aufreihen? Im Grunde ist diese Frage irrelevant, sie lässt sich aber als einfaches mathematisches Problem lösen.
Im Sinne der modernen Evolutionstheorie ist sie freilich sogar dumm gestellt. Denn sie geht davon aus, dass alle Buchstaben zum gleichen Zeitpunkt "auftauchten", nur in einer Anordnung "funktionieren" und nur in einem kulmulierten Moment, nicht nach einander oder in bestimmten vordefinierten Gesetzmäßigkeiten, aufgereiht werden.
Die viel wichtigere Frage ist, was notwendig ist, um eine Veränderung der Buchstabenkombinationen zu erreichen und wie lange es durchschnittlich dauert, bis es eine neue Buchstabenkombination gibt. Und erst dann ist zu überlegen, wie wahrscheinlich es ist, dass in einer bestimmten Zeit, bei einem rein zufälligen Verändern der Kombination, eine ganz bestimmte Buchstabenkombination entsteht.
Genau das, was die Evolutionstheorie beschreibt. Man muss dafür natürlich berücksichtigen, das es kein vordefiniertes Ziel gibt, der aktuelle Ist-Stand also auch hätte völlig anders sein können. Auch ist zu bemerken, dass der Evolution mit ungefähr 3 1/2 - 4 Milliarden Jahren ein unvorstellbar langer Zeitraum zur Verfügung stand und, dass es nicht lediglich ein Individuum gab, das hätte mutieren können.
Aus den Beobachtungen erschließt sich, dass die absolut überwiegende Anzahl an Mutationen entweder überhaupt keinen Effekt hat oder - und das ist schon wesentlich seltener - das Individuum nicht lebensfähig oder zumindest nicht lebensfähiger macht, als andere Individuen. Solche Mutationen werden durch eine gleichbleibende oder schlechtere Populationsrate ausgesiebt, also selektiert.
Bei der riesigen Anzahl an möglichen Mutationen und Individuen, die mutieren können, muss es auch Mutationen geben, die einem Individuum nützen. Das ist ebenfalls wieder nur eine Frage der Wahrscheinlichkeit. Diese Individuen haben durch eine höhere "Fitness" eine höhere Chance, ihr Erbgut in die nächste Generation weiterzugeben. Nützliche Mutationen werden also mit einer höheren Wahrscheinlichkeit weitergegeben. So gibt es in "kurzer" Zeit mehr Individuen, die bereits über diese Mutation verfügen.
Was allerdings nützlich ist, also die "Fitness" erhöht, bestimmt sich nicht objektiv, sondern allein danach, ob es einem Individuum in einer bestimmten Umwelt hilft oder nicht. In einer anderen Umwelt kann die gleiche Mutation fatal sein, was dazu führt, dass sie sich dort nicht durchsetzen kann. Und weil nützliche Mutationen relativ selten auftreten, ist Evolution ein eher langsamer Prozess, der nicht in den allerseltesten Fällen live beobachtet werden kann. Deshalb sehen Menschen, zum Beispiel, im Großen und Ganzen heute noch so aus, wie vor zehntausend Jahren. Aber ein paar tausend Jahre sind auch nichts im Vergleich zu 3 - 4 Milliarden Jahren.
Immer wieder liest man darüber hinaus ja auch - und im von mir rezensierten Video "Hat die Bibel doch Recht? - Der Evolutionstheorie fehlen die Beweise" wird es ebenfalls behauptet - dass es einen Widerspruch zwischen Darwin und Mendel gäbe.
Mendel untersuchte die reine Vererbung - ohne Mutation - von einer Generation auf die andere. Ausschlaggebend hierfür ist die Meiose.
Schema der Meiose. In diesem Beispiel sind drei Chromosomenpaare dargestellt und die Chromosomen je blau bzw. rot gekennzeichnet nach dem Elternteil, von dem sie geerbt wurden. Außerdem sind Mikrotubuli und Centrosomen (beide gelb-orange) dargestellt, um die Phasen der Teilungen besser unterscheiden zu können. Auf (1) Prophase I (hier dargestellt in der Unterphase der Diakinese), (2) Metaphase I, (3) Anaphase I, (4) Telophase I der ersten meiotischen Teilung folgt – nach einer hier nicht dargestellten Zwischenphase der Interkinese – die zweite meiotische Teilung mit (5) Prophase II, (6) Metaphase II, (7) Anaphase II, (8) Telophase II. Vier gleich große Zellen wie hier angedeutet entstehen beispielsweise bei der Spermatogenese, nicht aber bei der Oogenese.
Meiose (griech. "Verminderung", "Verkleinerung"), auch Reifeteilung genannt, ist eine besondere Art Kernteilung eukaryotischer Zellen (#1), die in zwei Schritten abläuft: Meiose I (#2) und Meiose II (#3). Dabei wird die Anzahl der Chromosomen halbiert und es entstehen genetisch voneinander verschiedene Zellkerne. Damit unterscheidet sich die Meiose grundlegend von der sonst gewöhnlichen Kernteilung, der Mitose. Diese lässt den Chromosomenbestand unverändert und bringt damit genetisch identische Zellkerne hervor.
Bei der Meiose wird aus einem doppelten di-poliden Chromosomensatz einen einfachen ha-poliden Chromosomensatz gemacht. Dieser findet sich in den Keimzellen der Eltern wieder, welche bei der Befruchtung miteinander verschmelzen. Erst dadurch wird ein neuer doppelter ha-polider Chromosomensatz geschmiedet, der dann zu einem doppelten di-poliden Chromomensatz ergänzt wird. Ausführlicher ist das alles in den Fußnoten 2 und 3 lesbar. Wenn das alles funktioniert, können dabei natürlich nur die Erbinformationen weitergegeben werden, die bisher in der Elterngeneration auch schon vorhanden waren. Das entspricht Mendels Lehre.
Und das sind ihre Grenzen. Sie stellt den Normalfall dar und gilt da nicht mehr, wo die Erbinformation verändert wird. Dies kann beispielsweise durch einen Fehler bei der Meiose oder aber auch durch Mutation geschehen. Es handelt sich hierbei also nicht um eine nach den mendelschen Regeln vererbte Eigenschaft und nur die Evolutionstheorie gibt weiteren Aufschluss.
Mendel bietet also lediglich die Möglichkeit der Erklärung, wie Merkmale von einer Generation in die nächste getragen werden. Die Vererbungslehre ist also sehr wohl Teil der synthetischen Evolutionstheorie und widerspricht ihr nicht.
#1 - Eukaryotischer Zellen
Eukaryoten oder Eukaryonten (Eukaryota) sind eine Domäne der Lebewesen, deren Zellen (Eucyten) einen echten Kern und eine reiche Kompartimentierung haben. Hierin unterscheiden sie sich von den beiden übrigen Domänen im System der Lebewesen, den prokaryotischen Bakterien und Archaeen mit procytischen Zellen.
Demnach sind Tiere, Pflanzen, Pilze, Algen, einige Bakterien und auch der Mensch Eukaryoten oder Eukaryonten.
#2 - Meiose I (Reduktionsteilung)
Prophase I
Die erste meiotische Teilung beginnt mit der Prophase I. Diese wird in fünf Stadien unterteilt:
- Im Leptotän beginnen die Chromosomen zu kondensieren. Bis zum Ende der Prophase I sind die Enden der Chromosomen, die Telomere, an der inneren Zellkernmembran befestigt. Jedes Chromosom besteht aus zwei identischen Chromatiden.
- Das Zygotän ist durch die Paarung der homologen Chromosomen gekennzeichnet, also die Aneinanderlagerung der von den beiden Eltern erhaltenen Exemplare eines Chromosomentyps. Diese exakte Chromosomenpaarung, auch Synapsis genannt, verläuft von den Enden her reißverschlussartig, indem sich zwischen beiden Chromosomensträngen der synaptonemale Komplex bildet, der beide Stränge zusammenhält.
- Im Pachytän kommt es zur weiteren Kondensation und die gepaarten Chromosomen bilden als Bivalent je eine aus vier Chromatiden bestehende Tetrade. In dieser Phase ereignet sich das Crossing-over, womit der Austausch homologer Chromatiden-Abschnitte zwischen Nicht-Schwesterchromatiden von gepaarten Chromosomen eingeleitet wird. Der synaptonemale Komplex zerfällt danach wieder.
- Im Diplotän zeigen sich daher die Paare zweier Chromosomen deutlich mit je voneinander abgesetzten doppelten Chromatiden. Nun fallen an den Tetraden als sogenannte Chiasmata jene Stellen auf, wo zwei der vier Chromatiden kreuzweise untereineinander verbunden sind, wenn denn ein Crossover stattgefunden hat.
- Mit der Diakinese endet die Prophase I, indem die Chromatidentetraden sich verkürzen, der Nucleolus sich auflöst, die Hülle des Zellkerns sich zerlegt und der Spindelapparat gebildet wird.
Metaphase I und Anaphase I
In der Metaphase I versammeln sich die gepaarten Chromosomen in der Äquatorialebene des Spindelapparats. Auch in dieser Phase können im Lichtmikroskop die Chiasmata sichtbar werden. In der anschließenden Anaphase I werden im Gegensatz zur mitotischen Anaphase nicht einzelne Chromatiden, sondern Chromatidenpaare zu den beiden Spindelpolen bewegt. Auf Grund des vorangegangenen Cross-overs sind die beiden zusammenhängenden Chromatiden jedoch nicht mehr identisch.
Telophase I
In der Telophase I liegt an jedem Pol dann jeweils nur noch ein Chromosom (mit zwei Chromatiden) jedes Typs vor. Es ist also zu einer Reduktion der Chromosomenzahl gekommen. Wie bei der mitotischen Telophase dekondensieren die Chromosomen nun und die Kernhülle bildet sich wieder. An die abgeschlossene Kernteilung schließt sich eine Zellteilung an. Der anschließende Zeitraum bis zum Beginn der zweiten mitotischen Teilung wird als Interkinese bezeichnet.
Bei einem menschlichen euploiden Chromosomensatz enthält der Zellkern der diploiden Zelle vor der meiotischen Teilung 23 Paare duplizierter Chromosomen, also 46 Chromosomen bzw. 92 Chromatiden. Nach der Meiose I hat jeder der beiden Tochterkerne 23 Chromosomen erhalten, die je aus einem Chromatidenpaar bestehen, also 46 Chromatiden.
#3 Meiose II (Äquationsteilung)
Im Anschluss an die Interkinese folgt die Meiose II. Sie ähnelt vom Ablauf her einer mitotischen Teilung, mit dem Unterschied, dass nur noch halb so viele Chromosomen vorliegen.
Prophase II
Die Chromosomen werden erneut kondensiert.
Metaphase II
Die noch aus zwei Chromatiden bestehenden Chromosomen werden in der Äquatorialebene angeordnet und am Centromer getrennt.
Anaphase II
Die getrennten Chromosomen werden einzeln den Tochterkernen zugeteilt.
Telophase II
Die Aufteilung ist abgeschlossen und die Kernhüllen bilden sich wieder aus.
Die aus der Telophase II hervorgehenden Zellkerne enthalten damit jeweils einen haploiden, unverdoppelten Chromosomensatz. Somit sind die vier Chromatiden einer jeden Tetrade der Prophase I jeweils zufällig auf vier verschiedene haploide Zellkerne verteilt worden.
Beide Vorgänge, Meiose I und Meiose II, zusammen bewirken eine effektive Durchmischung des Erbguts der beiden Eltern. Diese Rekombination erhöht die Zahl der genetischen Kombinationen innerhalb der Population und damit die Chance, dass in einigen Individuen für das Überleben günstige Merkmale zusammenkommen und so zu einem Selektionsvorteil führen. Im Vergleich zu asexuellen Organismen ermöglicht dies eine schnellere Evolution.